第7章 〜老化の進化的共通性〜
[目的]様々な種類の生物において、摂取カロリーを制限しながら飼育すると寿命が延長し老化関連疾患の進行を遅らせることができる。カロリー制限がどのような仕組みで老化を遅らせるのかを解明するための研究が行われた結果、サーチュイン(sirtuin)ファミリーというタンパク質が強く関連していることが分かった。そしてこのサーチュインを人工的に活性化することでカロリー制限を行うことなく、老化を抑制する効果を模倣することができるかもしれないのだ。アメリカではこの原理を用いた老化抑制薬の開発も現に行われている。酸化防止薬と並び次世代の老化抑制薬として爆発的なヒットを飛ばすことが予想される!
[単語説明]
Sir:silent information regulator(サーチュイン)、タンパク質の一種で酵素として働く。NADを加水分解すると同時にアセチル化されたヒストンからアセチル基を取り去る反応を触媒する。この結果ヒストンは不活性はクロマチン構造をとる。これはサイレンシングと呼ばれる、遺伝子の発現を抑制する効果をもたらす。
NAD:nicotinamide adenine dinucleotide、右下のような構造をしている。
NAD⁺の形で存在していて、高エネルギー電子2個とプロトン(H⁺)を受け取ってNADHとなりエネルギーを運搬する役割を果たす。細胞内でNAD⁺が多いとは、エネルギーが不足がちになっていることを意味している。よってカロリー制限を行うとNAD⁺が豊富になり、その結果サーチュイによるサイレンシングが起こり、老化を遅らせることになる。
インスリン/IGF-I・シグナル伝達系:インスリン受容体あるいはIGF-I受容体から始まるシグナルを伝達する経路のこと。Sir2がこのシグナル伝達経路に何らかの形で作用するこ
とで老化を遅らせていると考えられている。
IGF-I:insulin-like growth factor-I(インスリン様増殖因子-I)
ERC:extrachromosomal ribosomal DNA circle(染色体外に切り出された環状のDNA)、酵母のrDNA領域は100~200コピーのrDNAの単位が繰り返された構造になっている。よって相同組換えによって環状のrDNAが切り出される減少が起こる。これが蓄積して酵母の老化が起こると考えられている。
rDNA:ribosomal DNA(リボソームDNA)、リボソームとは細胞質中に遊離しているか、小胞体や核膜と結合して存在する小顆粒で、RNAをアミノ酸に変換してアミノ酸の連なったタンパク質を作る翻訳という作業を行う。このリボソームはリボソームRNA(rRNA)2本と50種類以上のタンパク質から成っている。rDNAとはこのrRNAをコードしているDNAのことである。
相同組換え:相同な塩基配列(塩基の並び方が同じということ)においていったんDNAが切断され、結合部分を入れ替えて再び結合すること。これによりDNAが組み込まれたり切り出されたりする。同じ塩基配列が繰り返されている領域で切り出しが起こると、切り出されたDNAは末端同士が結合し環状になる。
ヘテロクロマチン:染色体上の高度に凝縮された部分のこと。この領域にはほとんど遺伝子がなく、あったとしてもほとんど発現しないことが分かっている。
オーソログ:ある2種類の生物が持っている遺伝子が共通の祖先が持っていた1つの遺伝子に由来するものであったとき、それらをオーソログであるという。
7.1 酵母と線虫の老化のメカニズム
酵母というのはビールとかパンの製造に使われる菌のことで、線虫というのは体長1mmくらいの小さな線状の虫だ。二つとも人間とはかけ離れた生物だが、老化機構において進化的に保存されている部分もあるかもしれない。つまりこれらの生物で試してみて寿命が延びたとしたら、それを人間にも応用できる可能性があるのだ。
酵母は母細胞から娘細胞が出芽してきてある程度大きくなったら分裂する。それに比べ線虫の体細胞は全て分裂しない細胞(分裂停止細胞)からなっている。
表1 酵母と線虫の老化兆候
|
酵母
|
線虫
|
|
細胞表面の傷の蓄積
|
外的刺激に対する反応性の低下
|
|
細胞形態の変化
|
リポフスチン色素の蓄積
|
|
分裂時間の延長
|
体内器官の運動性・反応性の低下
|
|
接合能の消失
|
特定の酵素の活性低下
|
これらの老化による兆候は右表に示す通りである。このようにこれらはまったくかけ離れた生物であるが、Sir2と呼ばれる共通のタンパク質により老化や寿命が制御されていることが明らかになった。
注 リポフスチンとは脂質に富む黄褐色の色素粒子で人間では神経細胞や心筋細胞に蓄積し、老化を引き起こす原因になっていると言われている。
7.2 Sir2の効果
Sir2を欠く酵母は寿命が約半分まで短縮するのに対し、Sir2を1コピー余分に持つ酵母は寿命が30~40%延長し老化過程も遅くなる。酵母の老化は環状のrDNA(以下ERCと書く)が染色体から切り出され、母細胞の中に蓄積していくことで引き起こされる。染色体上のrDNA領域という同じ配列が繰り返されている部分から相同組み換えによりERCが切り出され、ERCが自己複製により母細胞の側に蓄積され、大量に蓄積されたERCが核小体の断裂化、膨化をもたらす。SIR2は酵母のrDNA領域における相同組換えを抑制しており、ERCの生成を抑制することによって、酵母の寿命を延ばしているのだ。(図1)Sir2は他のタンパク質とSir複合体を形成していて、接合遺伝子座(HML,HMR)、rDNA、テロメアに局在し、サイレンシング(遺伝子発現の不活化)を行っているが、老化に伴い接合遺伝子座、テロメアから核小体へと再局在化される。これにより母細胞は接合能を失い、最終的にはDNA複製や転写に必須の因子が大量のERCに奪われることで死ぬと考えられる。
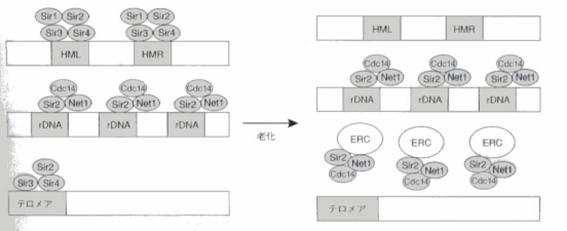
図1 老化した酵母におけるSirタンパクの再分配(文献2より引用)
線虫の場合Sir2オーソログであるsir-2.1を人工的に発現させると約50%寿命が延びる。このSir2の効果はインスリン/IGF-I・シグナル伝達系に深く関与していることが明らかになっている。またdaf-16はインスリン/IGF-I・シグナル伝達系の下流に位置する転写因子で、daf-2はインスリン/IGF-I・シグナル伝達系のはじめに位置するインスリン様因子受容体である。sir-2.1はdaf-16の上流でdaf-2からのシグナルに反応する遺伝子を制御しているか、daf-2のリガンドとなる因子あるいはその産生をコントロールする因子をコードする遺伝子を制御していると考えられる。

図2 線虫の老化(文献1より引用)
7.3 SIR2とは一体何なのか
Sir2とはNAD依存性ヒストン脱アセチル化酵素のことで、ヘテロクロマチン構造の構成成分としてSir3、Sir4などとともにSir複合体を形成し接合遺伝子座やテロメア領域における転写抑制(サイレンシング)を行ったり、別の複合体を形成してrDNA領域におけるサイレンシング及び相同組換えの抑制(これが老化の抑制に役立っている)を行っている。Sir2はNADの加水分解と基質の脱アセチル化反応を同時に行う酵素で、入手可能なNADの量が多いと脱アセチル化酵素を活性化し、クロマチンの構造構築を通してゲノムのサイレンシングを行う。つまり細胞のエネルギー状態をゲノムの制御状態へと変換する能力を持ったエネルギーセンサーとして機能していて、その結果老化、寿命を制御しているのである。

図3 Sir2のサイレンシング(文献1より引用)
別の言い方をすればSir2はNAD依存性脱アセチル化酵素としての活性を通してエネルギー代謝(特にグルコース代謝が重要である)と老化、寿命を関連づけしていると言える。酵母においては栄養分のグルコースを制限すると有意に寿命が延びるという事実はこれを証明している。
7.4 単純は動物と高等動物の共通性
ショウジョウバエというハエにおいてインスリン/IGF-I・シグナル伝達系が老化、寿命の制御に重要な役割を果たしているという報告がなされた。概略図におけるchico遺伝子(インスリン様因子受容体からのシグナルを下流のP13Kシグナル伝達系に伝達する)やInR(インスリン様因子受容体)遺伝子を変異させた個体は有意に寿命が延長した。マウスにおいても成長ホルモン-IGF-Iの系と老化、寿命が密接に関連していることが見出されている。酵母などの単細胞生物種においてはグルコースが細胞内のシグナル伝達に直接関わっていて、Sir2もそのシグナル伝達系の中で働く。線虫、ハエ、マウスなどの多細胞生物種においては、グルコース代謝は神経内分泌系から分泌されるホルモンによって調節され、細胞内ではインスリン/IGF-I・シグナル伝達系が重要な役割を担っている。つまり進化とともにSir2の作用の仕方も変わってきているようだ。
7.5 哺乳類の老化
酵母においてはSir2がゲノムの不安定性(ERCの産生)や不適切な遺伝子発現(接合能の消失)をNAD依存性脱アセチル化酵素として抑制し、老化・寿命を制御している。この構図が哺乳類においても成り立っているかもしれないということを示唆する事実が存在する。哺乳類のSir2オーソログがp53の転写活性化能を抑制することでアポトーシスを抑制しているということが明らかになった。P53はγ線や紫外線の照射、DNAに障害を与える薬剤、過酸化水素などによる酸化ストレスによってリン酸化、アセチル化を受け、転写因子として活性化されることが知られている。その結果、細胞増殖の停止やアポトーシスを引き起こす。マウスや人のSir2aはp53を脱アセチル化することで、p53の機能を抑制しているようだ。実際Sir2を強制発現させた細胞は上のダメージに対する抵抗力が上がるのである。

図4 酵母と哺乳類におけるSir2の機能(文献1より引用)
このSir2によるサイレンシングが環境的な要因で少しずつ崩壊していくと考えると、ダメージの蓄積によりそれまで発現していなかった遺伝子が機能し、老化の原因となるとともに、環境的要因の蓄積が同時に老化をもたらすと考えられる。つまり老化とはプログラム的な要素と確率的な要素を両方持っている現象であると言えるのである。
7.6 カロリー制限模倣化合物
これまでの理論から老化を遅らせる一つの手段はサーチュインを活性化する物質を見つけ出し、体内に取り込むことであることが分かる。サーチュイン活性化物質の活性の測定は試験管の中でp53の脱アセチル化を定量するという方法で行われる。こうして同定した物質はレスベラトロール、ブテイン、ピセアタンノールなどの植物性ポリフェノール化合物である。このうちレスベラトロールが最も強いサーチュイン活性を示す物質で、ブドウの皮や赤ワインに多く含まれているらしい。だが多くとれば良いというものでもないようだ。どの程度の量が最も効率的であるかはまだ分かっていない。



レスベラトロール ブテイン ピセアタンノール



イソリキリチゲニン フィセチン クエルセチン
図5 カロリー制限模倣化合物
7.7 これからの展望
これからの研究課題はサーチュイン活性化物質を設計し、医薬品として実用化していくことである。この研究により少なくとも老化の原因のうちの一つを克服し、人間の寿命を延長することができると期待できる。人類が若さを操るようになる時代もそう遠くはなさそうだ。もちろんこれは不老不死の実現までの長い過程のうちの一つのステップでしかないのだが。
(参考文献)
老化研究がわかる
実験医学 世界発 老化・寿命制御のメカニズム解明に挑む
不老不死への科学